
Fitness
Malassezia appears to facilitate Microsporum canis growth | IDR
Introduction
Tinea capitis is a superficial fungal infection that affected the scalp and hair, predominately occurring in children, particularly those between the ages of 2 and in 8. Infection is rare in adults.1 In children with tinea capitis, there is usually no significant difference in the proportion of affected boys and girls in. However, in adult cases of tinea capitis, female are more commonly affected.2,3 The main causative fungi for both children and adults are dermatophytes, which include anthropophilic, zoophilic and geophilic fungi.
In recent years, several studies have shown an increasing incidence of tinea capitis in healthy adults, which emphasizes the necessity for greater attention to this condition in future research.2,4 Adult tinea capitis often presents with mild inflammatory symptoms, making a challenge in differentiation from psoriasis capitis, seborrheic dermatitis and atopic dermatitis of the scalp.5 This might be attributed to the development of sebaceous glands during puberty, which secrete long-chain fatty acids known for their antifungal properties.2 It has been suggested that differences in scalp microorganisms between children and adults due to gradual maturation of the sebaceous gland during puberty, leading to increased sebum production and the lipophilic microorganisms such as Malassezia and Cutibacterium.6–8 After menopause, the incidence of tinea capitis tends to increase due to decreased sebaceous gland secretion and scalp oil.9
We encountered a 60-year-old female in our clinical setting with tinea capitis, exhibiting three types of fungal growth: Microsporum canis, Malassezia globose, and Malassezia furfur. Interestingly, we observed that Malassezia and Microsporum canis can coexist on the scalp, especially in regions with heightened sebum production. To investigate the interaction of Microsporum canis and Malassezia, we conducted in vitro co-cultures of isolated Microsporum canis with two species of Malassezia. Our finding indicated that Malassezia may promote the growth and activity of Microsporum canis. These findings hint at a potential association between the atypical manifestations of adult tinea capitis and specific Malassezia species.
Case Presentation
On May 1, 2022, a 60-year-old female presented to our department with complaints of rapid diffuse hair loss on the top of her scalp, accompanied by mild pain for a month. She had then been previously diagnosed with folliculitis of the scalp and received treatment with various antibiotics at a local hospital, but her symptoms did not improve. The patient has a history of type 2 diabetes, which has been well-managed with oral medications, including metformin, acarbose and glimepiride, with her blood glucose levels maintained within the normal range. There is no history of autoimmune diseases or tumors. Upon further investigation, it was revealed that the patient had been in contact with a dog before the onset of disease. However, no areas of shedding or rash were observed on the dog.
During the physical examination, patchily greasy erythema was observed on the top of the scalp, with fine flakes that were easily removable. The hair on the top of the head appeared sparse, with some strands being dry and tapered (Figure 1A). Wood lamp examination revealed bright green fluorescence in the area of hair loss on the top of the head (Figure 1B).
The patient’s blood routine and erythrocyte sedimentation rate examination were within normal limits. Fluoroscopy of the patient’s dandruff and hair showed a significant presence of mycelium and spores. The fungal culture results and molecular sequencing confirmed the coexistence of Microsporum canis, Malassezia globosa and Malassezia furfur.
The patient was diagnosed with tinea capitis based on the presence of a rash and findings from fluorescence microscopic. The patient was informed about the necessary precautions for managing conditions, including cutting her hair and disinfecting bedding sheets and other items. The patient was treated with oral itraconazole at a dose of 200 mg per day and topical ketoconazole ointment twice daily for a duration of 2 weeks. Prior to starting treatment, the patient had her hair cut short (Figure 1C). Three weeks later, the erythema on the top of the head had disappeared, desquamation was significantly reduced, the hair became thicker compared to before, and the fluorescence microscopy test showed a negative finding (Figure 1D). During the 2-month follow-up, there was no signs of erythema or desquamation, and no recurrence was observed at the 6-month- follow-up.
Mycological Study on the Causative Pathogen from Scalp Lesions
The Direct Observation of Causative Pathogen Under Fluorescent Microscopy and Light Microscopy
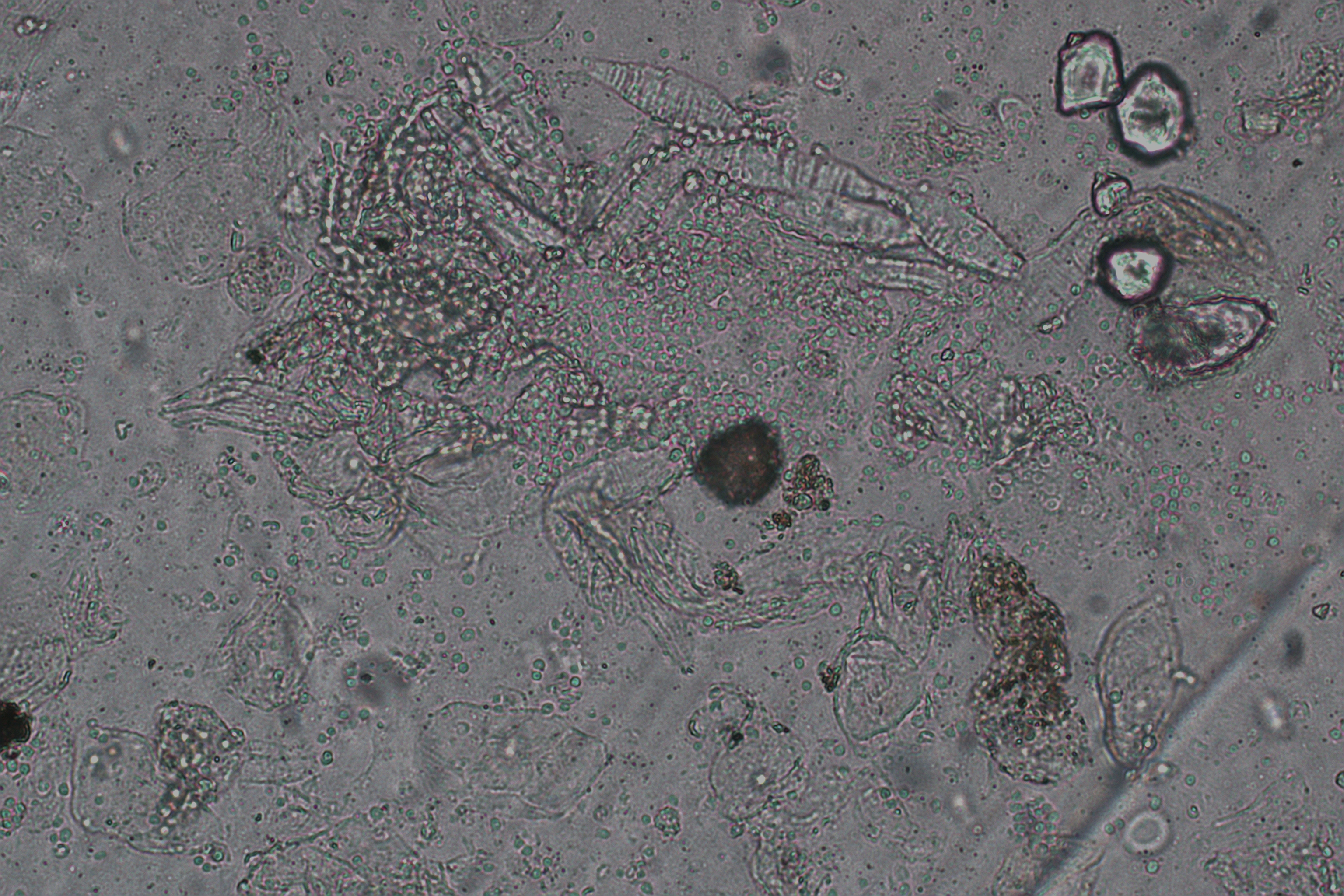
Specimens were collected for direct mycological examination. The scalp scales and hair were stained with calcofluor white stain (CFW) and examined under a fluorescence microscope. The results revealed the presence of antler-like, separated hyphae and globose spores (Figure 2A). Additionally, large spindle-shaped conidia were observed in the same field of view when examined under a light microscope (See Figure 2B and Supplementary Material 1). Further examination of scalp scrapings and hair from the affected area on the top of scalp showed a significant presence of antler branch hyphae and spherical spores both under a fluorescence microscope (Figure 2C) and a light microscope (Figure 2D).
{kind=link}
 |
Figure 2 Specimens from the skin lesion were subjected to direct microscopic examination. (A) a representative image of fungal hyphae and spores from the scalp scales on the top of head staining with calcofluor white under fluorescence microscope (original magnification, ×400); (B) represents the same field as A under the light microscopy. (C) a representative image for fungal hyphae and spores from hair in the rash area on the top of the head staining with calcofluor white under fluorescence microscope (original magnification, ×400); (D) represents the same field as C under the light microscope. (E) a representative image of fungal spores from the scalp scales on the occipital staining with calcofluor white under fluorescence microscope (original magnification, ×400); (F) represents the same field as (E) under the light microscopy. (G) a representative image for fungal spores from hair in the rash area on the occipital staining with calcofluor white under fluorescence microscope (original magnification, ×400); (H) represents the same field as G under the light microscope. |
In the occipital area, spherical spores of varying sizes were observed in the scalp debris under fluorescence microscope (Figure 2E), as well as in corresponding light microscope images (Figure 2F). Hair samples from this area showed the presence of attached spores but no hyphae when examined under a fluorescence microscope (Figure 2G) and a light microscope (Figure 2H). Generally, under direct microscopic examination, fungal load in dander and hair samples from the top of the scalp is greater than that observed in samples from the occipital area, as evidenced by more severe damage in the samples from the top of the scalp.
The Isolation and Identification of Causative Pathogen
Dander samples were scraped from the affected scalp area and the periphery, and then inoculated into a medium for a period of 7 days. The results revealed fast-growing colonies with villous-shaped mycelium on Sabouraud dextrose agar (SDA) from both the parietal region (Figure 3A) and occipital region (Figure 3B). Initially, the colonies appeared white at the center and gradually changed to grey towards the periphery. The cultured colonies from the top of the scalp samples grew faster compared to the occipital samples.
 |
Figure 3 The culture results of patients with tinea capitis. (A and B) the image for direct culture of colonies on Sabouraud’ s Dextrose agar (SDA) after sampling from the scales on the top of the head and occipital, respectively. (C and D) the image for direct culture of colonies on Modified Dixon agar after sampling from the scales on the top of the head and occipital, respectively. (E) a representative image for purification of colony of Microsporum canis after culture on Sabouraud’ s Dextrose agar at 37 °C for 1 week. (F) a representative image for purification of colony of Malassezia globosa after culture on Modified Dixon agar at 37°C for 1 week. (G) a representative image for purification of colony of Malassezia furfur after culture on Modified Dixon agar at 37 °C for 1 week. |
In addition, the colonies on mDixon agar displayed light yellow cheesy colonies and plush filamentous colonies, both from the parietal region (Figure 3C) and occipital region (Figure 3D). However, the filamentous colonies were more pronounced in the scalp samples, while the cheesy colonies were more pronounced in the occipital samples.
The primary cultured colonies from the parietal region were further purified on SDA and mDixon agar. Three colonies with different textures, shapes, and sizes were isolated. To identify the fungal strains, DNA sequencing was performed on the isolated strains using the ITS1/ITS4 region of ribosomal DNA (rDNA). The isolated fungi were identified as Microsporum canis (Figure 3E), Malassezia globosa (Figure 3F), and Malassezia furfur (Figure 3G).
To study the morphology and characteristics of Microsporum canis, the small-culture method was employed to cultivate and observe the characteristics. After 6 days of cultivation in the rice medium, a fluorescent dye was added to a slide. The cover slide was then removed from the petri dish and placed on the slide, allowing it to sit for 5 minutes before microscopic observation. Under both a light microscope (Figure 4A) and a fluorescence microscope (Figure 4B), the morphology of the microorganism was observed in the same field of view. The majority of the large conidia appeared spindle-shaped, with thick, rough, and prickly walls, as well as septal hyphae. However, the separation of mycelium and the morphology of the conidia were clearer and more accurate under the fluorescence microscope.
 |
Figure 4 Image showing microscopic observation of Microsporidium canis in the same part of the paddle after 7 days of rice medium culture. (A) Image showing Microsporidium canis under light microscope (original magnification, ×400); (B) Image showing Microsporidium canis staining with calcofluor white under fluorescence microscope (original magnification, ×400); (C) Image showing Microsporidium canis staining with lactophenol cotton lalue under light microscope (original magnification, ×400). |
Subsequently, lactophenol cotton blue dye was added from the edge of the same slide, and any excess dye was absorbed using filter paper on the other side. Once the entire slide was filled with lactophenol cotton blue dye, the morphology of Microsporum canis was observed under a light microscope in the same field of view as shown in Figure 4A and B. The results revealed clear peripheral contours of the mycelia and conidial morphology (Figure 4C).
The Co-Culture of Microsporum canis and Malassezia in vitro
To investigate whether Malassezia inhibits the growth of Microsporum canis, we conducted co-culture experiments using the isolated Microsporum canis and two species of Malassezia (Malassezia globosa and Malassezia furfur) from this patient.
First, both Malassezia globosa and Malassezia furfur were suspended in normal saline and diluted to a concentration of 1×104/mL. Then, 200 μL of each Malassezia suspension was added to individual plate containing SDA or mDixonA medium and subsequently spread evenly on the surface of the agar. Next, a 2×2 mm2 colony of Microsporum canis was picked up using a sterile pick stick and placed in the center of the medium. The growth of the Microsporum canis colony was observed. Control culture plates containing Microsporum canis or Malassezia only were also set up to monitor the colony growth.
The combination of these three fungal species led to the formation of five experimental groups: (1) Malassezia globosa group, (2) Malassezia globosa and Microsporum canis co-culture group, (3) Microsporum canis group, (4) Malassezia furfur and Microsporum canis co-culture group, and (5) Malassezia furfur group. All groups were cultured simultaneously in SDA and mDixonA media at temperatures of 27°C, 30°C, and 37°C.
As show in Figure 5A and B, after a 10-day culture period, Malassezia globosa did not growth in mDixonA medium at 27°C, but its growth increased at higher culture temperatures. Similarly, Malassezia furfur showed increased growth with higher temperatures. Conversely, in mDixonA medium, the diameter of Microsporum canis colonies decreased gradually at 27°C, 30°C, and 37°C. No growth of Malassezia was observed in SDA medium, and the diameter of Microsporum canis colonies decreased with increasing temperature (Figure 5C and D).
 |
Figure 5 The effect of Malassezia on the growth of Microspora canis in vitro. The experiment included groups: Microspora canis cultured alone, Microspora canis co-cultured with Malassezia globose, and Microspora Canis co-cultured with Malassezia furfur. The strains were inoculated on mDixon agar or Sabouraud dextrose agar (SDA) and incubated at temperatures of 27°C, 30°C, and 37°C for a period of 10 days. The colonies on mDixon agar (A) or SDA (C) were photographed and the diameter of the colonies was measured using the ImageJ software. The diameter of the colonies was measured and recorded on the Y-axis (B and D). To ensure reliability and consistency, the experiment was repeated three times. P is considered significant when +Malassezia furfur and Microsporum canis co-culture group VS Malassezia globosa and Microsporum canis co-culture group). |
Interestingly, was co-culturing Microspora canis with Malassezia furfur in the mDixon medium at 27°C, 30°C and 37°C resulted in larger colony diameter compared to when Microspora Canis was cultured alone (P= 0.0005, PPMalassezia globosa and Microspora Canis were co-cultured at 27°C and 30°C (P= 0.0005, P= 0.0251). However, no similar findings were found on SDA media.
These findings indicate that Malassezia and Microsporum canis can coexist in areas of the scalp with high sebum secretion. Additionally, Malassezia may even promote the growth and activity of Microsporum canis.
Discussion
Tinea capitis is primarily caused by tinea fungi, with Trichophyton and Microsporum species being the main culprits. In Europe and Southeastern Korea, Microsporum canis is still the most commonly reported causative agent of tinea capitis.4,10 In Portugal, the most common etiological agents are Microsporum audouinii (20%) and Trichophyton rubrum (20%).9 Changing in hygiene conditions and lifestyle in America and England, have resulted in a different epidemiology of dermatophytosis, with Trichophyton tonsurans emerging as the most prevalent species.11–13 A multicenter study has indicated that zoophilic agents are more likely to infect children in China, while adult patients are more susceptible to anthropophilic dermatophyte infections. Although the species spectrum of pathogenic fungi may vary across regions, Microsporum canis still remains the primary pathogen for both children and adult patients with tinea capitis.1
Lipophilic yeasts of the genus Malassezia are important skin commensals and opportunistic skin pathogens in a variety of animals. They live almost exclusively on the skin and mucosal sites of warm-blooded vertebrates14 and birds.15 The pathogenic potential of Malassezia for infection and its association with other diseases, such as cancer, have been increasingly reported.16–20 Malassezia species are the most abundant fungi found in all sampled healthy human skin body sites except the feet, with M. restricta and M. globosa standing out as the most common and most numerous. Microbiome dynamics revealed that healthy skin communities dominated by M. restricta, M. globose and M. furfur.21 Although the skin is the primary Malassezia reservoir, it can be found in other parts of the body, including the human and murine gut, human breast milk, and internal organs including those of the central nervous system.22 Various Malassezia species occur on human skin as commensals, and they are also associated with multiple skin disorders, such as pityriasis versicolor, Malassezia folliculitis, seborrheic dermatitis/dandruff, atopic dermatitis, and psoriasis.23
Recently, the incidence of tinea capitis in healthy adults is increasing, accompanied by notable changes in the microbial spectrum. These changes require warrant caution in future clinical practices. The alteration of the scalp microflora greatly influences the onset and progression of the disease, and the susceptibility of patients to tinea capitis may contribute to the shift in the scalp microbial environment.7,8,10 A study conducted by Tao et al observed that the Shannon diversity of the bacterial community was higher in tinea capitis patients when compared to the healthy scalp samples. Non-metric multidimensional scaling (NMDS) analysis at the genus level revealed interesting results. While two specimens exhibited the presence of Microsporum canis and Malassezia in culture, the NMDS results indicated a higher abundance of Malassezia compared to Microsporum canis, especially when Malassezia globosa and Malassezia restricta were presented.7
Tinea capitis is a rare condition in adults, with the main incidence occurring among postmenopausal women. Hormonal changes in the body after menopause, characterized by decreased blood estrogen levels, can contribute to this condition. The sebaceous glands are affected by these changes, leading to a reduction in the fungistatic action of long-chain fatty acids present in sebum.2,3,6 Additionally, the increasing prevalence of adult tinea capitis may be associated with underlying conditions such as diabetes, increased use of steroids and immunosuppressants, as well as reduced triglycerides with antifungal properties.5
In this case, we observed an adult patient presenting with seborrheic dermatitis in the scalp rash area, accompanied by hair loss. A significant number of fungal spores were found in the affected area, and the sample under direct microscope observation and culture results in mDixon medium confirmed the presence of Malassezia spp. Malassezia tends to flourish and multiple in areas of the human body where sebum secretion is abundant due to the absence of fatty acid synthetase.24 The skin lesions of our patient contained a significant amount of lipid components, providing an ideal environment for the growth of Malassezia. Interestingly, the direct observation under microscope and fungal culture results revealed the coexistence of Microsporum canis, Malassezia globosa, and Malassezia furfur in the lesion area, particularly in regions with high sebum secretion. It is worth noting that the clinical manifestations of tinea capitis in adults often differ significantly from those in children and may present with atypical rashes resembling seborrheic dermatitis.25 Our findings suggest that Malassezia may have a promoting role in the development of tinea capitis, which could explain the atypical clinical manifestations observed in adult patients.6
Jo et al observed that prepubertal skin hosts a variety of fungi, while Malassezia species dominate adult skin, and the fungal community on the skin undergoes significant changes during puberty.8,26 During puberty, as sebaceous glands differentiate and mature, sebum secretion increases, which is associated with a decrease in tinea capitis incidence. Conversely, after menopause, sebaceous gland secretion decreases, leading to an increase in tinea capitis incidence.27 Although previous studies have shown that ethyl ester derivatives generated through the esterification of medium-chain fatty acids by Malassezia exhibit in vitro antimycotic activity,28 and Malassezia globosa secretes aspartyl protease 1 (MgSAP1) with demonstrated in vitro antibacterial properties.29 Our co-culture experiments yielded opposite results. We observed that Malassezia may play a role in promoting the growth of Microsporum canis. This could be attributed to the potential consumption of long-chain fatty acids by Malassezia, which weakens its antibacterial effect and may contribute to the occurrence of adult tinea capitis.27,30 These findings suggest that tinea capitis rashes may not be caused by a single strain and may be related to the balance of microorganisms in the scalp environment. Malassezia, as the most common commensal flora, especially on the scalp, warrants greater attention whenever addressing skin conditions.
Although the growth of Microsporum canis was increased by the presence of Malassezia in vitro, the pathogenic role of Malassezia in this case remains to be determined. We recognize the limitations inherent in this study, as it stands, this report represents a singular case study, necessitating additional cases to substantiate our hypothesis regarding the interaction between different fungal species.
In summary, our case report and co-culture experiment suggest a potential promoting role of Malassezia in the development of adult tinea capitis. While Malassezia is responsible for the manifestation of seborrheic dermatitis in the scalp, it may contribute to conditions like tinea capitis by consuming long-chain fatty acids. Further research is necessary to explore the complex relationship among skin microbiome and understand the mechanisms involved in interaction between different fungal species. To provide more accurate guidance for clinical diagnosis and treatment, it is crucial for future investigations and experiments to monitor the pathogenic impact of microbiome changes.
Data Sharing Statement
All data generated or analyzed during this study are included in this published article.
Ethics Approval and Informed Consent
This study was approved by the Institutional Research and Ethics Committee of Jining No. 1 People’s Hospital to publish the case details (Ethical approval no. 2022-029). The patient provided written informed consent for publication of this case report and any accompanying images. The study was carried out in accordance with the principles of the Declaration of Helsinki. The first author vouches for the completeness and accuracy of the data and for the adherence of the study to the protocol.
Consent for Publication
Written informed consent was obtained from the patient described in this report. A copy of the written consent is available by request.
Acknowledgments
We thank the patient for granting permission to publish this information.
Funding
This work was supported in part by grants from the National Natural Science Foundation of China (NM 82272358), Key Research and Development Plan of Jining (2023YXNS001), Medicine health science and technology development plan of Shandong Province (202202070556).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Chen XQ, Zheng DY, Xiao YY, et al. Aetiology of tinea capitis in China: a multicentre prospective study. Br J Dermatol. 2022;186(4):705–712. doi:10.1111/bjd.20875
2. Liang G, Zheng X, Song G, et al. Adult tinea capitis in China: a retrospective analysis from 2000 to 2019. Mycoses. 2020;63(8):876–888. doi:10.1111/myc.13102
3. Toksöz L, Güzel AB, Ilkit M, Akar T, Saraçlı MA. Scalp dermatophyte carriage in pregnant, pre-, and postmenopausal women: a comparative study using the hairbrush and cytobrush methods of sample collection. Mycopathologia. 2011;171(5):339–344. doi:10.1007/s11046-010-9377-3
4. Lee HJ, Kim JY, Park KD, et al. Analysis of adult patients with tinea capitis in Southeastern Korea. Ann Dermatol. 2020;32(2):109–114. doi:10.5021/ad.2020.32.2.109
5. Mayser P, Nenoff P, Reinel D, et al. S1 guidelines: tinea capitis. J Dtsch Dermatol Ges. 2020;18(2):161–179.
6. Park J, Schwardt NH, Jo JH, et al. Shifts in the Skin Bacterial and Fungal Communities of Healthy Children Transitioning through Puberty. J Invest Dermatol. 2022;142(1):212–219. doi:10.1016/j.jid.2021.04.034
7. Tao R, Zhu P, Zhou Y, et al. Altered skin fungal and bacterial community compositions in tinea capitis. Mycoses. 2022;65(8):834–840. doi:10.1111/myc.13480
8. Jo JH, Deming C, Kennedy EA, et al. Diverse human skin fungal communities in children converge in adulthood. J Invest Dermatol. 2016;136(12):2356–2363. doi:10.1016/j.jid.2016.05.130
9. Duarte B, Galhardas C, Cabete J. Adult tinea capitis and tinea barbae in a tertiary Portuguese hospital: a 11-year audit. Mycoses. 2019;62(11):1079–1083. doi:10.1111/myc.12991
10. Polak-Witka K, Constantinou A, Schwarzer R, et al. Identification of anti-microbial peptides and traces of microbial DNA in infrainfundibular compartments of human scalp terminal hair follicles. Eur J Dermatol. 2021;31(1):22–31. doi:10.1684/ejd.2020.3948
11. Zhi H, Shen H, Zhong Y, et al. Tinea capitis in children: a single-institution retrospective review from 2011 to 2019. Mycoses. 2021;64(5):550–554. doi:10.1111/myc.13243
12. Ginter-Hanselmayer G, Weger W, Ilkit M, Smolle J. Epidemiology of tinea capitis in Europe: current state and changing patterns. Mycoses. 2007;50(Suppl 2):6–13. doi:10.1111/j.1439-0507.2007.01424.x
13. Nguyen CV, Collier S, Merten AH, Maguiness SM, Hook KP. Tinea capitis: a single-institution retrospective review from 2010 to 2015. Pediatr Dermatol. 2020;37(2):305–310. doi:10.1111/pde.14092
14. Lorch JM, Palmer JM, Vanderwolf KJ, et al. Malassezia vespertilionis sp. nov.: a new cold-tolerant species of yeast isolated from bats. Persoonia. 2018;41(1):56–70. doi:10.3767/persoonia.2018.41.04
15. Guillot J, Bond R. Malassezia yeasts in veterinary dermatology: an updated overview. Front Cell Infect Microbiol. 2020;10:79. doi:10.3389/fcimb.2020.00079
16. White SD, Vandenabeele SI, Drazenovich NL, Foley JE. Malassezia species isolated from the intermammary and preputial fossa areas of horses. J Vet Intern Med. 2006;20(2):395–398. doi:10.1111/j.1939-1676.2006.tb02874.x
17. Pin D. Seborrhoeic dermatitis in a goat due to Malassezia pachydermatis. Vet Dermatol. 2004;15(1):53–56. doi:10.1111/j.1365-3164.2004.00369.x
18. Dinsdale JR, Rest JR. Yeast infection in ferrets. Vet Rec. 1995;137(25):647–648.
19. Pinter L, Anthony RM, Glumac N, Hajsig D, Pogacnik M, Drobnic-Kosorok M. Apparent cross-infection with a single strain of Malassezia pachydermatis on a pig farm. Acta Vet Hung. 2002;50(2):151–156. doi:10.1556/avet.50.2002.2.3
20. Kuttin ES, Glas I. Mycotic otitis externa in animals. Mykosen. 1985;28(2):61–68. doi:10.1111/j.1439-0507.1985.tb02097.x
21. Proctor DM, Dangana T, Sexton DJ, et al. Integrated genomic, epidemiologic investigation of Candida auris skin colonization in a skilled nursing facility. Nat Med. 2021;27(8):1401–1409. doi:10.1038/s41591-021-01383-w
22. Spatz M, Richard ML. Overview of the potential role of malassezia in gut health and disease. Front Cell Infect Microbiol. 2020;10:201. doi:10.3389/fcimb.2020.00201
23. Prohic A, Jovovic Sadikovic T, Krupalija-Fazlic M, Kuskunovic-Vlahovljak S. Malassezia species in healthy skin and in dermatological conditions. Int J Dermatol. 2016;55(5):494–504. doi:10.1111/ijd.13116
24. Theelen B, Cafarchia C, Gaitanis G, Bassukas ID, Boekhout T, Dawson TL. Malassezia ecology, pathophysiology, and treatment. Med Mycol. 2018;56(suppl_1):S10–S25. doi:10.1093/mmy/myx134
25. Yang Z, Chen W, Wan Z, Song Y, Li R. Tinea capitis by microsporum canis in an elderly female with extensive dermatophyte infection. Mycopathologia. 2021;186(2):299–305. doi:10.1007/s11046-020-00519-9
26. Lin Q, Panchamukhi A, Li P, et al. Malassezia and Staphylococcus dominate scalp microbiome for seborrheic dermatitis. Bioprocess Biosyst Eng. 2021;44(5):965–975. doi:10.1007/s00449-020-02333-5
27. Buckley DA, Fuller LC, Higgins EM, du Vivier AW. Lesson of the week. Tinea capitis in adults. BMJ. 2000;320(7246):1389–1390. doi:10.1136/bmj.320.7246.1389
28. Mayser P. Medium chain fatty acid ethyl esters – activation of antimicrobial effects by Malassezia enzymes. Mycoses. 2015;58(4):215–219. doi:10.1111/myc.12300
29. Li H, Goh BN, Teh WK, et al. Skin commensal Malassezia globosa secreted protease attenuates Staphylococcus aureus biofilm formation. J Invest Dermatol. 2018;138(5):1137–1145. doi:10.1016/j.jid.2017.11.034
30. Wu G, Zhao H, Li C, et al. Genus-wide comparative genomics of malassezia delineates its phylogeny, physiology, and niche adaptation on human skin. PLoS Genet. 2015;11(11):e1005614.

From Stadium to Screen: How Technology is Changing Sports Viewing

20/7/2024 Horse Racing Tips and Best Bets – Flemington, Flemington Cup day

Indian tech hub Karnataka state’s move to reserve jobs for locals not finalised, chief minister says

Financial picture dramatically improves for Shamrock Rovers in 48 hours with Sinclair Armstrong deal and European win

Our fashion editor’s favourite affordable bag is 20% off today

Jay Shah’s Big Decision Hovers Over Cricket’s Associate Member Directors Election

Budget shampoo that adds MAJOR volume boost is on sale for Amazon Prime Day: ‘Your hair feels fuller and thicker’

Being active on your commute lowers risk of disease and mental health

Tiger Woods cops brutal Open schedule as full Round 1 tee times revealed
